

Eine neue Interleukin 10-Reportermaus
BLOG: Die Sankore Schriften

![]()
Du spazierst nachts durch die Straßen einer Reihenhaussiedlung. Als Du an einem roten Backsteinhaus vorbeikommst, geht plötzlich das Licht an dessen Haustür an. Der Bewegungsmelder hat dein Kommen bemerkt und automatisch die Hausbeleuchtung angeschaltet. Die Hausbewohner im Innern wissen nun, dass sich da draußen jemand dem Haus genähert hat. Wenn Forscher die Regulation der Genexpression in lebenden Zellen beobachten wollen, befinden sich in einer ähnlichen Situation wie die Hausbewohner, die nichts sehen können, sondern nur indirekt über den Bewegungsmelder und das Licht erfahren, das draußen vor der Haustür etwas geschieht.
Um etwas über die Expression eines bestimmten Gens zu erfahren, benutzen die Wissenschaftler deshalb einen genetischen Bewegungsmelder der Ihnen sagt wann und wo dieses Gen aktiv ist. Diese genetischen Bewegungsmelder nennt man Reportersysteme. Genau, wie der Bewegungsmelder am Haus das Licht anschaltet, schalten diese molekularen Sensoren in der Zelle ein Licht an, was man dann im Mikroskop oder entsprechende fluoreszenzdetektierende Geräte sehen kann. Dieses Licht wird durch die Expression eines sogenannten Reportergens erzeugt.
Zurück zu unserem Nachtspaziergang: Stell Dir mal vor, dass statt eines erwachsenen Mannes von 1.80 m eine Maus oder gar zehn Mäuse am Eingang des Hauses vorbeiflitzen. Würde die Hausbeleuchtung angehen? Bestimmt nicht. Die Maus ist zu klein und zu schnell für den Bewegungsmelder.
Wollte man also mit einem herkömmlichen Bewegungsmelder entdecken, ob sich nachts Mäuse am Hauseingang tummeln und ob es wenige oder viele sind, stünde man vor einer schwierigen Aufgabe. Genau vor so einer großen Herausforderung stand der Immunologe Hicham Bouabe vom Max von Pettenkofer-Institut in München, als er mithilfe einer Reportermaus untersuchen wollte, welche der verschiedenen Immunzellen in der lebenden Maus Interleukin 10 (Il-10) exprimieren.
Was tat Bouabe also? Erstens erfand einen neuen Bewegungsmelder, der sehr viel empfindlicher war als die herkömmlichen, sodass dieser nun auch Mäuse entdecken konnte und zweitens stattete er das Licht mit einem Dimmer aus. Das hieß: Es leuchtete umso stärker, je mehr Mäuse der Bewegungsmelder entdeckte [3].
Das war der erste Teil, indem es darum ging, die Idee von Bouabe zu veranschaulichen. Im zweiten Teil dieses Blogbeitrags werde ich nun für die Genetiker und solche die es werden wollen einige technische Details seines innovativen Reportersystems beleuchten.
Doch bevor es ans Eingemachte geht, werde ich euch erstmal erzählen, warum es so wichtig ist, sich mit der Expression von Il-10 in verschiedenen Zelltypen zu beschäftigen. Aus Platzgründen werde ich nur einige Beispiele von Vielen herausgreifen: Il-10 wurde erstmals 1989 als ein Faktor entdeckt, der von Maus TH2-Zellen produziert wird und die Zytokinproduktion von TH1-Zellen inhibiert. Aufgrund dieser Wirkung wurde es als Cytokine Synthesis Inhibiting Factor (CSIF) bezeichnet. Mittlerweile ist jedoch bekannt, dass nicht nur TH2-Zellen, sondern weitere unterschiedliche Zelltypen, wie Makrophagen, Dendritische Zellen, Mastzellen, TH1-Zellen, B-Zellen etc., IL-10 produzieren können. Im Vergleich zu anderen Zytokinen wie z. B. TNF-α ist die zelluläre Produktion von IL-10 aber relativ schwach und seine Halbwertszeit beträgt 60-180 Minuten. Das offene Leseraster von IL-10 codiert für ein 178 Aminosäuren großes Vorläuferprotein, das eine Signalsequenz zur Steuerung des Transports vom IL-10-Protein enthält. Das reife IL-10, das schließlich ausgeschüttet wird, wird seiner Signalsequenz entledigt, und ist nur noch 160 Aminosäuren lang.
IL-10 spielt eine zentrale Rolle bei der Deaktivierung mehrerer entzündungsvermittelnder Signalwege und der Dämpfung bzw. Inhibierung der Aktivität vieler Immunzellen. Seine immunsuppressive Schlüsselrolle zeigt sich u.a. beim LPS-induzierten septischen Schock. Die Vorbehandlung von Mäusen mit rekombinanten IL-10 führt zu deren Schutz bei letaler LPS-Gabe. Weiterhin entwickeln IL-10 defiziente Mäuse eine überschießende T-Helfer-1 (TH1)-Antwort und erkranken spontan an einer chronischen Enterocolitis, einer der Colitis ulcerosa beim Menschen ähnlichen Darmerkrankung, und stellen damit ein wichtiges Versuchsmodell für chronisch entzündlichen Darmerkrankungen (CED) dar.
Interessanterweise scheinen diverse Krankheitserreger die immunsuppressive Funktion von IL-10 zu ihren Gunsten zu nutzen, indem sie dessen Expression induzieren, um dem Immunsystem zu entkommen. Legionella pneumophila, Salmonella typhimurium, Mycobacterium tuberculosis, Mycobacterium avium, Mycobacterium leprae, Listeria monocytogenes, HIV-1, Rhinovirus, Respiratory syncytial virus (RSV) und Yersinia enterocolitica sind nur einige Beispiele solcher Erreger, die während ihrer Infektion eine starke IL-10-Expression induzieren können. Vor diesem Hintergrund ist es notwendig, die entscheidenden pathophysiologisch relevanten Zelltypen für IL-10-Produktion herauszufinden.
Bouabe baute zunächst eine IL-10-Reportermaus bei der die Translation von EGFP, einem grün fluoreszierenden Protein, durch eine interne Ribosomen-Eintrittsstelle (IRES) an die von IL-10 gekoppelt war [2]. IRES erlaubt eine CAP-unabhängige Bindung der Ribosomen und Initiierung der Translation. Es entstand also eine bicistronische mRNA für Il-10 und EFGP. In der nächsten Abbildung seht ihr eine schematische Darstellung dieses Reportersystems
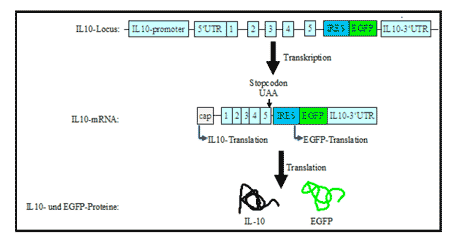
Abb. 1: Schematische Darstellung der Transkription/Translation des IL10-IRES-EGFPReportersystems Herstellung einer IL10-IRESEGFP-Reportermaus, in der IRESEGFP-Sequenz hinter dem Stoppcodon und vor der 3’nichttranslatierenden Region eingebaut ist. Nach der Transkription wird eine sogenannte Cap-Struktur (7-Methyl-guanosin) am 5’Ende der mRNA angeheftet. An der Cap-Struktur binden Translation-Initiationsfaktoren, welche die Rekrutierung von Ribosomen und die Translation von IL-10 einleiten. Die Translation des ersten Cistron endet am IL-10-Stopcodon. Die Ribosomen werden auch an die interne Ribosomen-Eintrittsstelle (IRES) rekrutiert und leiten, unabhängig von der Cap-Struktur, die Translation von EGFP ein. Die Kästchen mit den Zahlen 1 bis 5 repräsentieren die 5 Exons von IL-10. 5’UTR und 3’UTR: 5’ bzw. 3’ nicht-translatierte Region.
Um zu prüfen, ob die EGFP-Fluoreszenz im FACS detektierbar ist, wurden CD4-T-Zellen aus der Milz der Reportermäuse isoliert und anschließend in der Zellkultur für 3 Tage z.B. mit einem speziellen CD28-Antikörper (genannt Superagonist), D665, stimuliert. Dendritische Zellen und Makrophagen, welche aus dem Knochenmark dieser Reportermäuse stammten, wurden 36 Stunden mit 100 ng/ml LPS stimuliert. In CD4-T-Zellen konnte die EGFP-Fluoreszenz detektiert werden. Sie entsprach dabei der IL10-Produktion im Kulturüberstand. Dendritische Zellen und Makrophagen aus dieser Reportermaus zeigten keine messbare EGFP-Fluoreszenz, obwohl sie eindeutig IL-10 produzierten. Das wurde mittels des Cytometric Bead Array (CBA) nachgewiesen.
Durchschnittlich sind 1×105 EGFP-Moleküle nötig, um deren Fluoreszenz im FACS detektieren zu können. Je stärker die Granularität der Zellen (z. B. bei Makrophagen, Dendritische Zellen, Granulozyten), desto mehr EGFP-Moleküle sind nötig, damit die Reporteraktivität sich von der Autofluoreszenz der Zellen abhebt. Damit entgehen solche und andere granulären Zellen, sowie Zellen mit schwacher IL-10-Produktion, der Erfassung mittels dieses IL10-Reportermausmodells. Dies bedeutet, dass dieses IL-10-EGFP Mausmodell nicht für die IL-10 Expressionsanalyse nutzbar ist. Eine weitere Einschränkung der Sensitivität dieser Reportermaus liegt in der Translation von EGFP über die IRES-Sequenz. IRES-abhängige Translation ist um ca. 50-80% weniger effizient als die Cap-abhängige Translation.
Aus den dargelegten Gründen hat Bouabe systematisch daran gearbeitet, Alternativlösungen zur Erhöhung der Reporteraktivität zu finden. Zunächst testete er die Möglichkeit, ob über ein IRES-vermitteltes polycistronisches Reportersystem eine Erhöhung der Reporteraktivität erzielt werden kann. Die Antwort war ja! [1] Zweitens suchte er nach einem alternativen sensitiveren Reporterprotein. Hier wusste ich, dass nur ein Enzym als Reporter infrage kommen kann, teilte Bouabe mit. Denn während jedes EGFP-Molekül nur einen Fluorophor bildet, kann jedes Enzymmolekül viele fluorogene Substrat-Moleküle umsetzen und damit Fluorophore in großem Maßstab erzeugen.
Allerdings sollte dieses Reporterenzym, so Bouabe, noch folgende Bedingungen erfüllen:
a) Das Enzym und sein Substrat dürfen nicht toxisch für die Zellen sein.
b) Das Substrat sollte membrandurchlässig sein, um mit intakten Zellen arbeiten zu können.
c) Das Produkt der Substratumsetzung sollte hoch polar sein, damit es nicht aus der Zelle diffundiert.
d) Die enzymatische Aktivität sollte einfach in einzelnen intakten Zellen detektierbar sein.
Bouabe fand, dass nur ein Enzym all diese Eigenschaften erfüllte: das Enzym TEM1-ß-Lactamase (Bla). Entscheidend für seine Wahl war das Vorhandensein eines für Bla geeignetes fluoreszierendes Substrat, genannt CCF2-AM (oder CCF4-AM), welches im Jahr 1998 von der Arbeitsgruppe des Nobelpreisträgers Roger Y. Tsien entwickelt wurde.
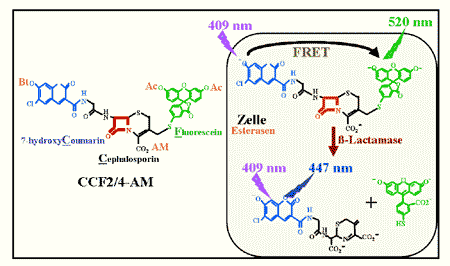
Abb. 2: Die Bla-Substrate CCF2/4-AM CCF2-AM bzw. CCF4-AM bestehen aus zwei Fluorophoren Hydroxycoumarin und Fluorescein, die über ein Cephalosporin-Molekül miteinander verbunden sind. Die Ester-Verbindungen (Ac:Acetyl; Bt: Butyryl; AM: Acetoxymethyl) neutralisieren die Ladungen des Substrats und machen es dadurch membrandurchlässig. Nach der Diffusion ins Zellinnere katalysieren zytoplasmatische Esterasen die Hydrolyse der Esterbindungen. Wegen seiner nun poly-anionischen Form, wird CCF2/4 in der Zelle zurückgehalten. Im intakten Substrat führt die Anregung von Coumarin (Donor) bei 409 nm zu einem Fluoreszenz Resonanz Energie Transfer (FRET) auf Flourescein (Akzeptor). Dadurch wird Fluorescein angeregt und strahlt grünes Licht aus (detektierbar bei 520 nm). Die ß-Lactamase katalysiert die Hydrolyse des Substrats, was zur Trennung vom Donor (Coumarin) und Akzeptor (Flourescein), und damit zur Unterbrechung des FRET, führt. Die Anregung des Donors (Coumarin) führt dann zur Emission vom blauen Licht (detektierbar bei 447 nm).
Die veresterte Form von CCF2/4, genannt CCF2/4-AM (AM: Acetoxymethyl), ist lipophil und kann somit leicht Zellmembranen passieren. Einmal ins Zellinnere gelangt, werden die vier Esterbindungen des Substrats durch unspezifische zytoplasmatische Esterasen hydrolysiert. Es entstehen negative Ladungen, wodurch die Rückdiffusion von CCF2/4 nach außen verhindert wird. Da das Substrat grün und das Produkt blau fluoreszieren, kann durch die Bildung des Verhältnisses zwischen der blauen Emission bei 447 nm und der grünen Emission bei 520 nm die Menge des umgesetzten Substrats, unabhängig vom Zelltyp, der Zellgröße, der Zellzahl oder der aufgenommenen Substratmenge, ermittelt werden. Im Gegensatz zu EGFP, das einige Stunden zur „Reifung“ benötigt bevor man es detektieren kann, kann die Expressionsmenge von ß-Lactamase in Echtzeit verfolgt werden. Veränderung im Emissionsverhältnis des Substrats kann schon bei der Expression von 50 ß-Lactamase- Molekülen gemessen werden (zum Vergleich, sind mindestens 1×105 EGFP-Moleküle notwendig um g Fluoreszenz zu detektieren). Darüber hinaus hat ß-Lactamase eine Halbwertszeit von ca. 206 Minuten und ähnelt damit der von IL-10 (ca. 60-180 min). Dies erlaubt die Durchführung von kinetischen Studien.
Um bei dem sehr schwach exprimierten IL-10 eine höchstmögliche Reportersensitivität erreichen zu können, hat Bouabe sein entwickeltes polycistronisches Reportersystem mit der ß-Lactamase als Reportergen kombiniert und eine IL10-2xIRES-Bla-Reporter (ITIB)-Maus generiert.

Abb. 3: Das 2xIRES Bla-Reportersystem der ITIB-Maus
Den Praxistest bestand diese ITIB-Reportermaus mit Bravour. Mit ihr konnte nämlich u.a. gezeigt werden, dass F4/80+ Makrophagen die Hauptquelle von IL-10 in verschiedenen Organen von naiven Mäusen sind. Ferner konnte mithilfe der ITIB-Maus gezeigt werden, dass Neutrophile Hauptproduzenten von IL-10 während bakterieller Infektionen sind. Neutrophile sind die ersten Immunzellen, die nach einer Infektion ins Gewebe einwandern, und die Eindringlinge abtöten. Die ITIB-Maus ermöglicht uns auch kinetische Studien auf Einzelzellniveau in allen myeloiden und lymphoiden Zelltypen. Zusätzlich kann sie zum In Vivo Screening für zelltypspezifische Modulatoren der IL-10 Expression eingesetzt werden. Dies könnte von großer Bedeutung für die Pharmaindustrie sein, die an der Entwicklung von Wirkstoffen, die die IL-10-Produktion ankurbeln oder inhibieren sollen, forschen.
Weiterführende Literatur
Bildnachweis
Abb.1: Schematische Darstellung der Transkription/Translation des IL10-IRES-EGFPReportersystems
Quelle: Aus [2] Abb. 3.7. S. 83
Abb. 2: Die Bla-Substrate CCF2/4-AM
Quelle: Aus [2] Abb. 3.23. S. 103
Abb.3: Das 2xIRES Bla-Reportersystem der ITIB-Maus
Quelle: Aus [3] Fig. 2A S. 5
Bouabe, H., Y. Liu, M. Moser, M. Bösl and J. Heesemann (2011). Novel Highly Sensitive IL-10–ß-Lactamase Reporter Mouse Reveals Cells of the Innate Immune System as Substantial Source of IL-10 In Vivo J. Immunol.