

(Dietrich Starck und) das Kopfproblem (II)
BLOG: Anatomisches Allerlei

Im ersten Teil ging es um die Enstehung des "Kopfproblems" aus dem Geiste der Osteologie, der (vergleichenden) Knochenlehre. Wir haben gesehen, dass einige – aber keineswegs alle – Knochen im Zentrum und an der Basis des Schädels ontogenetisch – indem sie aus Knorpelvorstufen hervorgehen – tatsächlich Wirbeln ähneln. Der grössere Teil des Schädels jedoch – praktisch der ganze Gesichtsschädel und das Schädeldach – lässt sich nicht aus Wirbeln ableiten. Diese Knochen entstehen direkt im Bindegewebe. Die Osteologie ist also – was das Kopfproblem angeht – zweideutig. Aber jetzt tritt die Neuroanatomie auf den Plan.
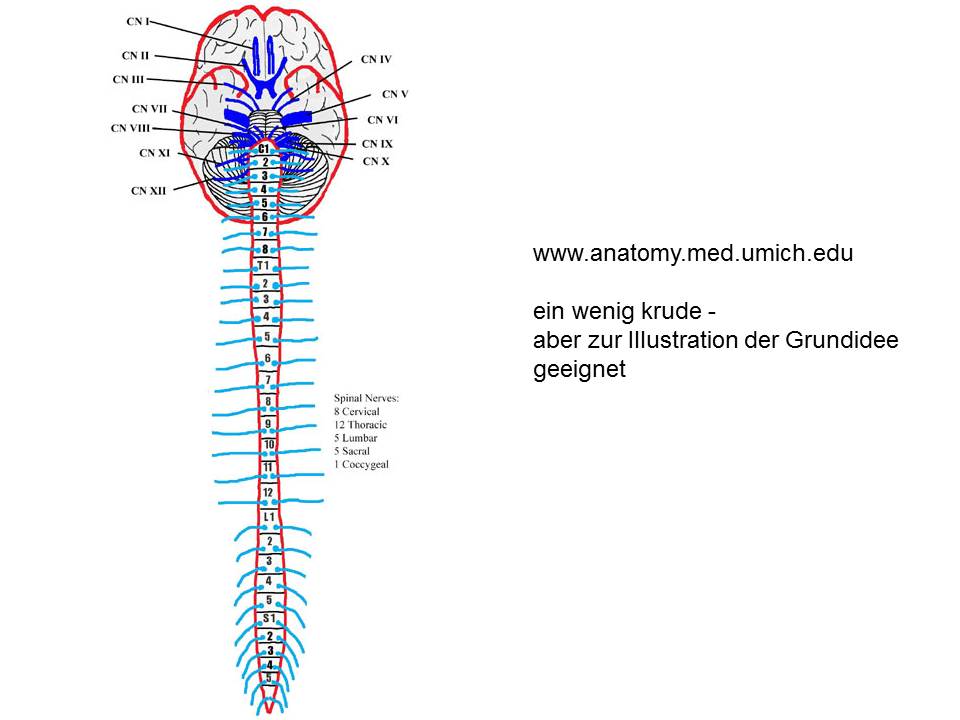
Wenn Kopf und Rumpf aus einer gemeinsamen Struktur hervorgingen, dann müssen die Nerven, die das Hirn mit Kopf und Hals und die das Rückenmark mit dem Rumpf verbinden, homolog sein. Seriell homolog. Auf den ersten Blick sind sie’s auch – Strippen, die rechts und links, seriell gereiht, aus dem Nervensystem hervorkommen.
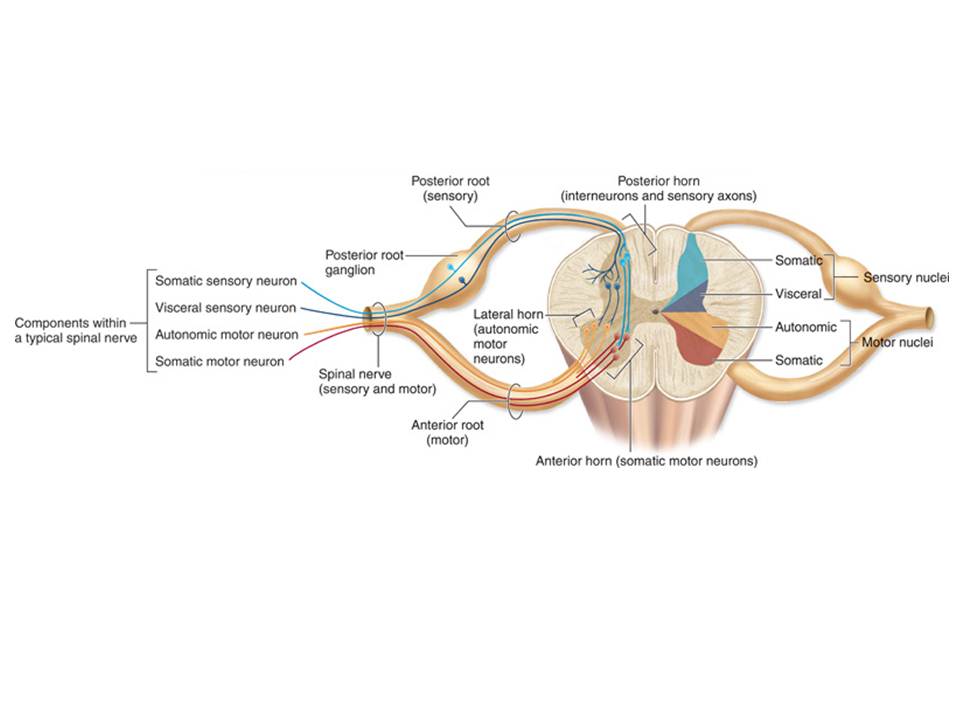
Auf den zweiten Blick dann wieder nicht. Denn – das lehren wir unsere Studenten – ein Rückenmarksnerv besteht aus einer ganglionbewehrten Radix dorsalis, die im Foramen intervertebrale mit der glatten Radix ventralis verschmilzt. Erstere, so lehren wir seit Bell und Magendie, ist rein sensorisch, die zweitere rein motorisch.

Das klappt im Kopf nun ganz und gar nicht. Denn es gibt zwar Nerven mit Ganglien und solche ohne, aber sie verschmelzen an den Austrittspunkten aus dem Schädel nie zu etwas, was einem Spinalnerv ähnelte. Ausweg: man vergleiche nicht einen einzelnen Hirnnerven mit einem einzelnen Spinalnerven, sondern mit jeweils einer Radix des Spinalnerven. Man postuliert, mit anderen Worten, dass einzelne Hirnnerven den nicht-fusionierten Radices der Spinalnerven entsprächen. Der ganglienbewehrte Trigeminus zum Beispiel wird dann zum Homologon einer dorsalen Wurzel und man muss, um zu einem kompletten Spinalnerven zu kommen, ihm einen anderen, aganglionären Hirnnerven zuordnen, der dann die ventrale Radix dieses Kopfsegmentes repräsentiert.

Das ganze gipfelt dann in Schemata wie diesem, die auf den ersten Blick harmlos aussehen, auf den zweiten Blick aber alles am Kopf in ein segmentales Schema zwingen und eine ungeheuere Menge an Daten integrieren Die Kästchen t-h sind präotische Somiten, die schraffierten Balken Branchiomeren. Das dritte hab’ ich farbig hervorgehoben, es ist der Mandibularbogen, den Sie alle kennen. Davor soll es allerdings noch zwei geben, mehr oder weniger hypothetische … das sind die vorhin erwähnten pro- und praepromandibulären Branchiomeren. Jedem Branchiomer wird artig ein Somit und ein Nervenpärchen zugeordnet, wobei – schauen wir uns wieder das dritte an – der Mandibularis des Trigeminus der dorsalen Wurzel dieses Segments entsprechen soll, die ventrale Wurzel dieses Segments aber durch gleich zwei Nerven repräsentiert wird, nämlich den allbekannten Trochlearis und den weniger populären Nervus rarus (nr), der bei Quastenflossern den basicranialen Muskel innerviert, der auch nicht gerade allzu bekannt ist. Der Nervus tenuis (nt) – auch er ist eine ein wenig obskure Struktur, die nur vorübergehend bei einigen Salamandern auftritt – er wird gebraucht, damit das Segment des Glossopharyngeus zu einer ventralen Wurzel kommt.
Sie sehen schon – das wird sehr schnell sehr verzwickt.
Aber es ist noch verzwickter, den eigentlich hätten Sie – eingedenk des Bell-Magendie’schen Gesetzes und der Wasserdichtigkeit der Physikumsfragen – schon längst entsetzt aufschreien sollen. Wie kann man denn einen unter anderem motorischen Nerven wie den Trigeminus oder den Glossopharyngeus mit einer rein sensorischen Hinterwurzel eines Spinalnerven vergleichen? Doch, man kann. Man ruft die vergleichende Anatomie zu Hilfe, die einem aber gleich auch wieder einige Beine stellt. Aber ich wollt’ ja mal einen Gedankengang aus dem Umfeld des Kopfproblemes in seiner ganzen Verzwicktheit darstellen. Also:
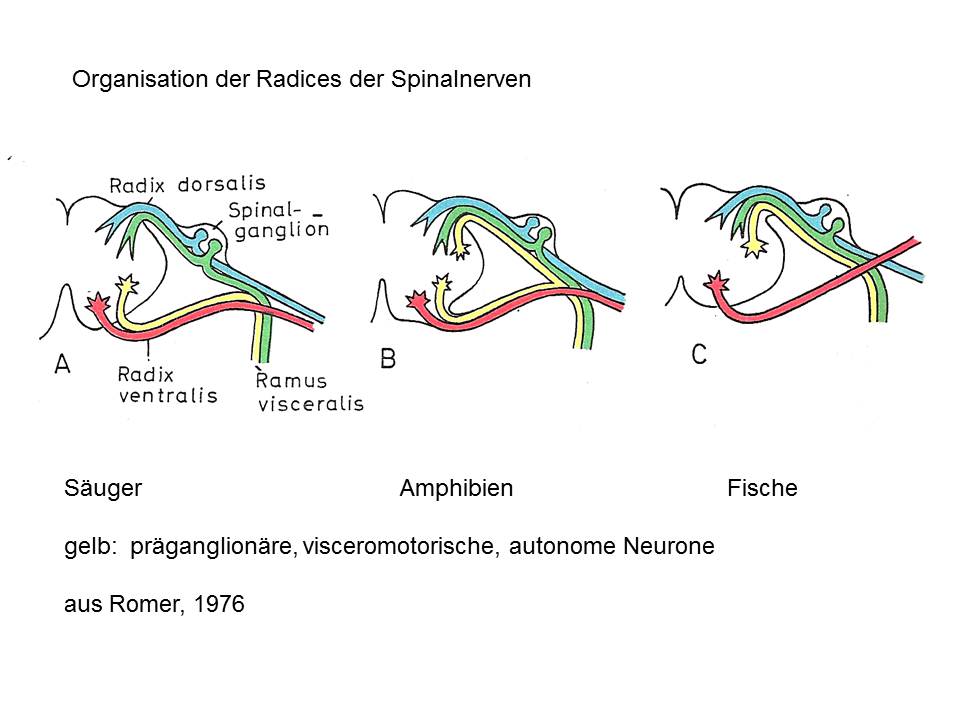
Dass die Hinterwurzel rein sensorisch sei, ist ein Lusus naturae, der die Säugetiere betrifft. Anderswo – bei Amphibien zum Beispiel – gibt es stets ein paar visceromotorische, präganglionäre, autonome Fasern in der Radix dorsalis. Bei vielen Fischen liegen diese präganglionären Fasern alle in der Radix dorsalis. Also, Hypothese: die Hinterwurzel ist ursprünglich gemischt sensomotorisch, der Vergleich, die Homologie zu einem ganglientragenden Hirnnerven ist also zulässig.
Die vergleichende Anatomie, dergestalt zur Hilfe gerufen, erweist sich also als freundliche Magd des Homologisierens, nur um gleich darauf wieder zum Advocatus diaboli zu mutieren. Denn, bittschön, was wird hier denn behauptet? Doch, dass die visceromotorische, autonome, präganglionäre Komponente, die sich in manchen Spinalnerven findet, derjenigen Motorik, die sich in Trigeminus und Glossopharyngeus findet, homolog sei.
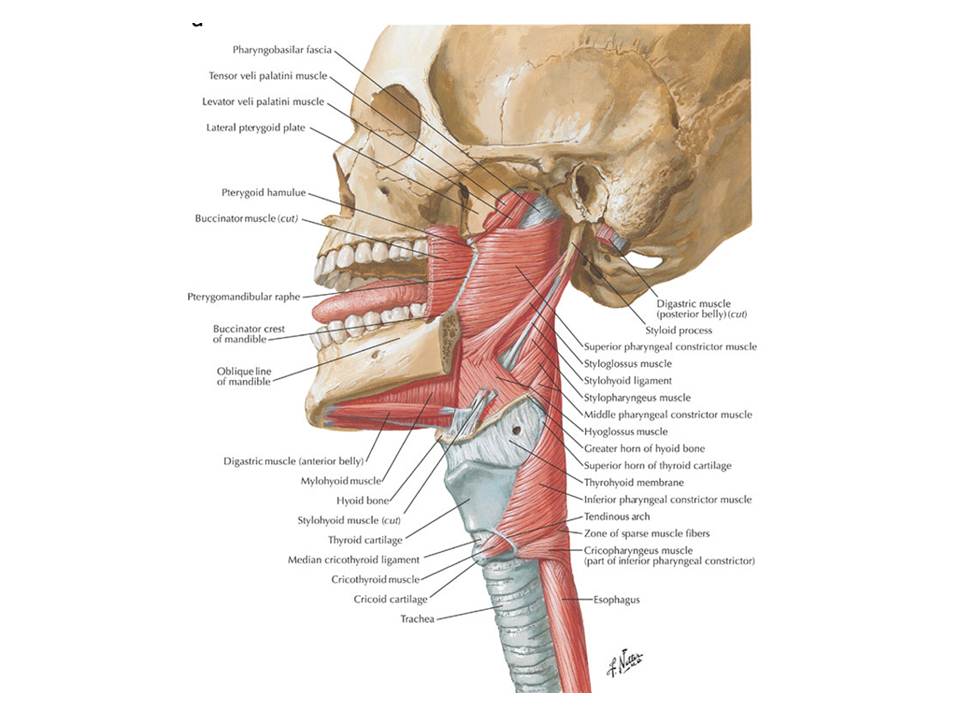
Und hier – gnadenlos – lehrt uns die Anatomie das Gegenteil. Es sind quergestreifte Skelettmuskeln, Kau- und Schluckmuskeln, die jene Hirnnerven innervieren. Die Muskeln sind nicht glatt wie jene des Darmes, um die sich die Visceromotorik sorgt. Keine Spur von Neuronenketten, wie sie für das visceromotorische System des Rumpfes typisch sind. Keine Spur von parakriner Transmission am Zielorgan: ganz handfeste neuromuskuläre Endplatten zieren die Enden der Axone. Diese Nerven sind eher somato- als visceromotorisch.
Nun, so hiess es, um die Idee zu retten, ursprünglich sei wohl auch da vorne am Kopfdarm die Muskulatur glatt gewesen und die Innervation sei über Neuronenketten erfolgt, dann, mit der Spezialisierung des Kau- und Schluckapparates, habe sich das alles geändert. Und ausserdem möge man bedenken, dass die Muskulatur, um die es ginge, ja immerhin in der Darmwand läge, wie am Rumpf, und wohl auch aus Seitenplattenmesoderm hervorginge…
und man bescherte uns dieses Wortungetüm, das heute noch die Lehrbuchautoren, die Dozenten der Anatomie (gar nicht zu reden von den Studenten) in den Wahnsinn treibt:
Dia "spezielle Visceromotorik"
Ich weiss nicht, wie es Ihnen geht, ich krieg’ nervöse Ausschläge, wenn ich das höre. Das ist kein Terminus technicus, das ist ein Desiderat, der wortgewordene Wunsch, Dinge vergleichen zu können, die man nicht vergleichen kann. Die Motorik des Kopf- und des Rumpfdarmes eben, nur, damit man die Nerven ebenfalls vergleichen kann. Mittlerweile sind übrigens auch noch die letzten Argumente, die sich zur Beibehaltung dieses Terminus vorbringen liessen, hinfällig geworden. Die Muskulatur, um die es geht, liegt zwar tatsächlich in der Kiemendarmwand – der Ausdruck "Branchiomotorik" wäre also allemal besser – entsteht aber nicht dort, entsteht nicht aus dem visceralen Blatt des Seitenplattenmesoderms, sondern aus Somiten. Wenn schon, dann also "spezielle Somatomotorik". Aber das würde, ebenso wie der Begriff der "Branchiomotorik", einer seriellen Homologie von Hirn- und Rückenmarksnerven den Boden unter den Füssen wegziehen.
Graben wir noch etwas tiefer, gehen wir in noch mehr Details. Wie lässt sich die Idee, dass einzelne Hirnnerven einzelnen, nicht-fusioniert Radices von Spinalnerven entsprächen, überhaupt begründen?
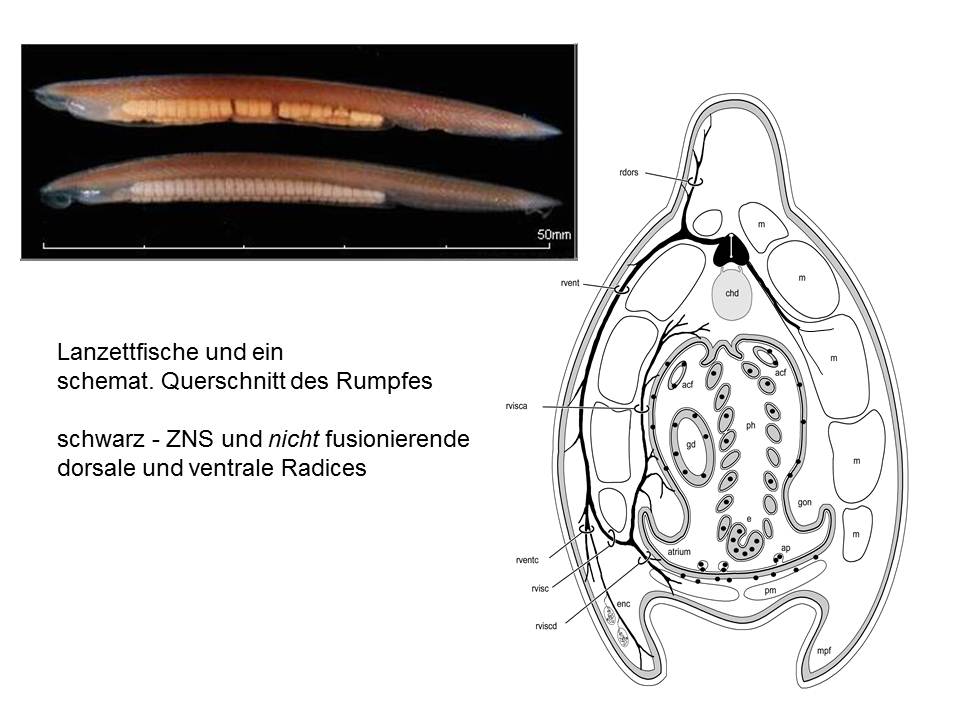
Der kleine Organismus, an dem ich momentan arbeite, der Lanzettfisch, liefert zunächst hochwillkommene Argumentationshilfe. Auch er hat ein Rückenmark, auch er hat dorsale und ventrale Radices, die daraus hervorgehen: aber sie fusionieren nicht. Bei Neunaugen ist es übrigens genauso. Seht, hiess es: das ist der Beweis! Die Nicht-Fusion der Radices ist das ursprüngliche Merkmal, das am Kopf erhalten geblieben ist, die Fusion am Rumpf ist das abgeleitete Merkmal!
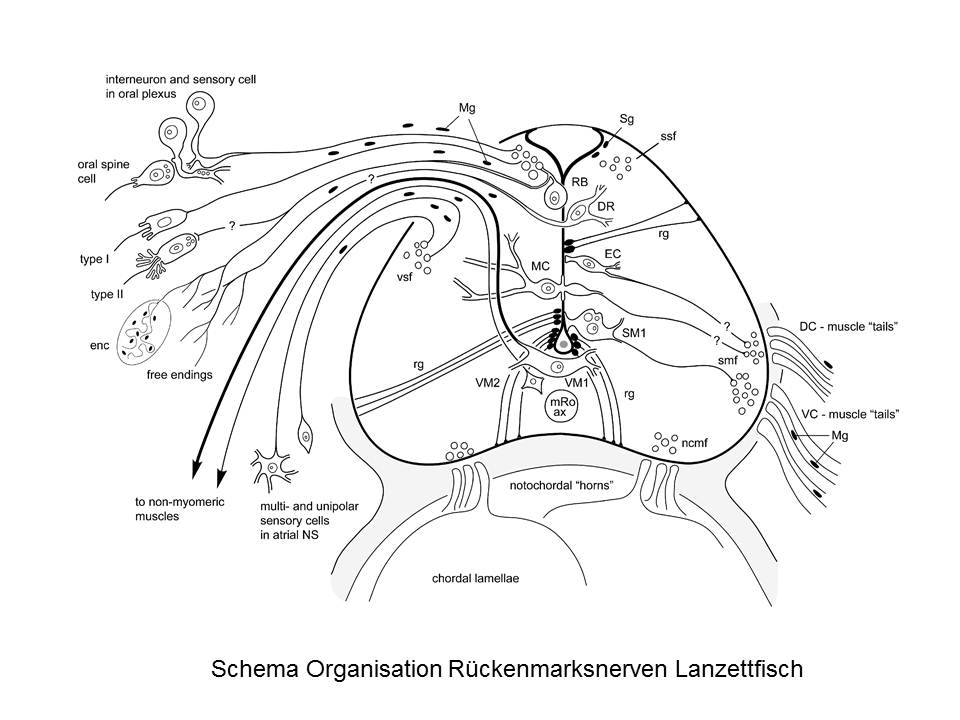
Alles wunderbar? Nein. Denn der Kronzeuge wackelt, wenn man ihm genauer auf den Zahn fühlt. Die ventralen Radices sind nämlich gar keine, es sind vielmehr Ausläufer ("muscle tails") der quergestreiften Muskelfasern selbst, die sich die ihre Innervation am Rückenmark sozusagen abholen.
Und die dorsalen Radices? Nun, auch sie sehen ganz anders aus, als jene eines Wirbeltiers. Es fehlt ihnen das Ganglion – dafür gibt es jede Menge intramedullärer sensorischer Neurone. Sie bestehen ausserdem zum grossen Teil aus den Axonen primärer Sinneszellen in Haut und Eingeweiden, die bei den Wirbeltieren auch weitgehend fehlen. Und – ja, sie enthalten eine Komponente, die man als "visceromotorisch" bezeichnet – aber die innerviert quergestreifte Muskeln. De facto besitzt das ganze Tierchen vermutlich gar keine glatte Muskulatur!
Sie sehen – anstatt den Lanzettfisch zum Kronzeugen der Verteidigung aufzurufen. kann man ihn ebensogut zu dem der Anklage machen. Die Basis des Vergleiches wackelt.
Das war der versprochene Ausflug in eines, nur eines der Labyrinthe des Kopfproblems. Antworten habe ich Ihnen keine geben können, aber hoffentlich ein Gefühl dafür, wie es im Inneren des Problems zugeht.
Im folgenden – und letzten Teil – wird es um Dietrich Starck gehen. Und um die Hox-Gene.
Meine Güte und ich tue mir schwer am Lig
Meine Güte, onomatopoiesis in Vollendung. Sollten Sie doch mal Hoffman Alberts weises Wasser versuchen oder einen Besuch in der Mexikanischen Wüste nebst Botanischem Experiment in erwägung ziehen, möchte ich dabei sein.
Ich dachte schon, ich hätte etwas mitbekommen aber Sie haben mich nur gestreift.
sascha von annosellemols
Persönlich finde ich den Ductus Boticelli nicht gefährlich für einen al.med.dent.
Schade, dass jemandem, der soviel zu sagen und keinen zum Zuhören hat nicht das Schweigen gelehrt werden konnte.
Dennoch sind auch bedauerliche und traurige Momente stets ein Zugewinn.
Einstein-Rosen-Brücke – komme ich mit meiner schweren Kugel aus Wissen den Berg raufgerollt, zu Dir, denn verstehen macht den Arzt
Vergleichende Anatomie?!
Erst einmal vielen Dank für diese drei Interessanten Artikel! Aber es stellt sich mir da eine Frage:
Gibt es denn kein Lebewesen oder Fossil das einen evolutionär unvollendeten Schädel oder WS hat? Sozusagen ein Mittelstück? Der Schleimaal hat doch meines Wissens nach Knorpelanlagen die einem Schädel/WS zumindest ähneln. Kann daraus nichts weiter geschlossen werden?
Gibt es viell ein Tier wo die Wirbelstruktur am Cranium noch erkennbar ist?
Mit freundlichen Grüßen
K.H.
@ K.H.
Das geht jetzt an’s Eingemachte:
Die Pars basilaris des Os occipitale entsteht tatsächlich aus somitischem Material, das weiter caudal Wirbel bildet. Bei manchen Fischen – Stören, wenn ich das recht entsinne – werden sogar einige primär zunächst voneinander getrennte “Hals”wirbel (was ist der “Hals” eines Fisches?)von caudal her mit dem eigentlichen Schädel verschmolzen.
Weiter cranial aber – siehe Text – ist von einem “Wirbelursprung” der Kopfknochen weit und breit nicht zu sehen.
Neunaugen und Schleimaale machen keine Ausnahmen von dieser Regel – ärger noch: die haben noch nicht mal Wirbel. Und noch eins “weiter unten” im Kladogramm, bei den Lanzettfischen, gibt es gar kein Kopfskelett.